Ongoing projects
Streptophyte algae and early plant evolution
Streptophyte algae and the evolution of land plant stress response
Embryophytes have evolved strategies to cope with the challenges of a terrestrial lifestyle. At the heart of these adaptations are land plants’ stress signaling networks. But what about the earliest steps of plants on land? These must have been enabled by exaptations that were present in the closest algal relatives of land plants—the streptophyte algae. To investigate the stress signaling capacities of land plants’ closest algal relatives, we challenged one representative species of each of the six major classes of streptophyte algae—including Zygnema circumcarinatum, a Zygnematophyceae and hence the class that has been pinpointed to represent the closest algal relatives of land plants—to the terrestrial stressors cold and high light.
Our recently published (see de Vries et al. 2018) differential gene expression analysis revealed pronounced changes in photosystem maintenance and other plastid biology-associated processes—as observed when challenging land plants with these plastid physiology-impairing cues. Of all six streptophyte algae investigated, Zygnema circumcarinatum invested the highest transcript budget to plastid biology-associated gene expression, representing an intriguing finding in light of our recent work (see below).
Importantly, we found many stress signaling circuits known from land plants to be responsive across the analyzed streptophyte algae. Importantly, Zygnema circumcarinatum was distinct by having a gene orthologous to the canonical abscisic acid receptors of the PYR/PYL family. This putative ABA receptor now awaits further investigation. Yet, regardless of the functionality, this has important implications for the earliest land plants’ genetic potential (see our recent review, de Vries and Archibald, 2018, on the genetic potential in streptophyte algae) for evolving stress signalling capacities. In sum, our data highlight that land plant textbook stress signaling gene networks were present in their algal ancestors, likely providing aid during streptophyte terrestrialization.
The evolution of nuclear control over plastid function
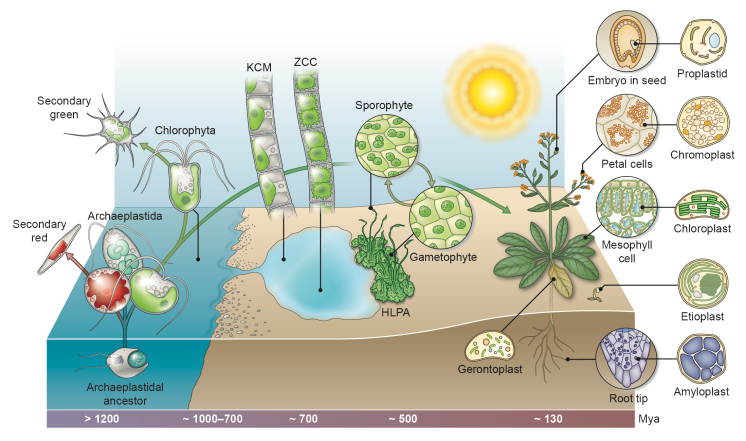
The origin of land plants was a singular event. Of all the photosynthetic eukaryotes, only one particular lineage conquered land on a global scale and we know that it belonged to the streptophyte algae. But which streptophyte algal lineage was it and can we tell why only this was so successful? Concerning the first question we know that it must have been a lineage sharing a close ancestry with one of the higher branching streptophyte algae, the Zygnematophyceae, Coloechaetophyceae and Charophyceae – which we coined ZCC (see de Vries et al. 2016).
Phylogenomics suggests that the Zygnematophyceae are the closest streptophyte algal relatives to land plants. Interestingly, presence/absence patterns of plastid-encoded proteins (and hence phylogeny-independent data) highlight Zygnematophyceae as having those plastids that are most similar to those of land plants (cf. de Vries et al. 2016). Moreover, these patterns draw a distinct line between plastids of lower branching and plastids of higher branching streptophyte algae; the latter having been inherited by the land plants. This warrants attention, as it might be relevant for one of the big questions in early plant evolution: what factors fostered the evolution of plants from that one particular lineage of streptophyte algae?
Streptophyte algae are thought to bear exaptations that have promoted terrestrialization. Among these is the ability to tolerate the terrestrial stressors high light and drought. Interestingly, both of these stressors are intertwined with plastid function and stress signaling fostered by plastid-nucleus communication. What if plastid properties and plastid-nucleus communication were part of these exaptations? I am currently exploring this idea by analyzing global gene expression patterns of streptophyte algae cultivated under conditions that induce an elevated plastid-nucleus communication.
Streptophyte plastid evolution
Plastid-encoded proteins can be very useful markers in streptophyte evolution. Two particularly interesting proteins are FtsH and YCF1. FtsH was introduced into plastid-bearing eukaryotes through the endosymbiotic acquisition of the cyanobacterial plastid progenitor (i.e. it was encoded in that ancient cyanobacterial genome). While it got transferred to the nucleus through endosymbiotic gene transfer, a copy stayed in the plastid genomes of red and green algae. Yet, in green algae plastid-encoded FtsH is often distinctly longer than its nuclear, rhodoplast-encoded or cyanobacterial homologs. Along the trajectory of streptophyte evolution, FtsH changed into YCF2, a protein of unknown function. This change from FtsH to YCF2 occurred within streptophyte algal evolution. Encoding YCF2 unites the plastids of higher branching streptophyte algae with those of land plants.
A similar pattern can be observed for another plastid-encoded protein of unknown function, YCF1. In de Vries et al. 2015 we described that along the trajectory of streptophyte algal evolution, YCF1 changed from a chlorophyte-specific type (which we coined Cte-YCF1) into a type shared by higher branching streptophyte algae and land plants (Ste-YCF1). Indeed, we later synthesized that changes in FtsH and YCF1 seemed to have happened at the same point of streptophyte evolution (de Vries et al. 2016). With the new wealth of sequence data on streptophyte algae, there are undoubtedly more of these patterns to be discovered.
For more details see de Vries et al. 2013, 2015, 2016, and 2017.

Previous projects
Plastid-stealing sea slugs
For recent developments on the topic click here or here
Biology text books listing differences between animal and plant cells exclaim that only plants have plastids (chloroplasts). Sacoglossan sea slugs do not follow this rule. Unlike plants, however, they do not inherit those plastids. Every generation of slugs has to steal them anew. Plastid acquisition occurs during the feeding process, when the sacoglossans prey on their siphonaceous macroalgal food. During that process, an unknown mechanism leads to the specific retention of only the plastids, while all other cellular content of the algae gets digested.
Depending on the slug species, stolen plastids (coined kleptoplasts) can be retained for up to several months. During that time, the plastids can show high levels of photosystem II activity. The main question that researchers working on the slugs have pondered is: how is that possible. Only the plastids but not the algal nuclei are retained by the slugs. The animal hence cannot synthesize those proteins that plants and algae use for chaperoning their plastids. For a long time it was thought that the slugs had acquired the genes necessary for plastid maintenance via lateral gene transfer. Yet, these genes proved to be undetectable in large-scale sequencing analyses, including ours – for a recent summary of our position on this, see Rauch et al. (2015). Moreover, global expression analysis carried out as part of my PhD project showed that starving slugs respond very little towards plastid status (cf. de Vries et al. 2015). Instead of focussing on the slugs, the plastid genomes caught our attention. What if the longevity of the plastids was actually due to plastid intrinsic properties. Algal plastids, in contrast to those of land plants, encode the proteins TufA and FtsH. The former being necessary for ongoing protein biosynthesis and the latter being an integral part of the photosystem II repair cycle. Especially FtsH hence warrants attention with regards to plastid longevity. We proposed that encoding FtsH equips the stolen plastids with the means to repair their photosystem II in a nucleus-independent manner – keeping also photooxidative stress at bay. Experimental testing of this idea is currently carried out in a collaboration between MolEvol and the Nickelsen lab at the Ludwig Maximilian University of Munich.
